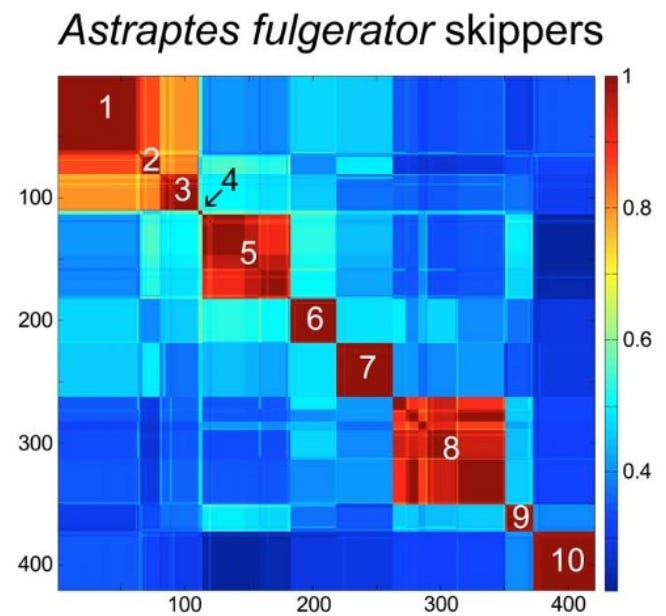
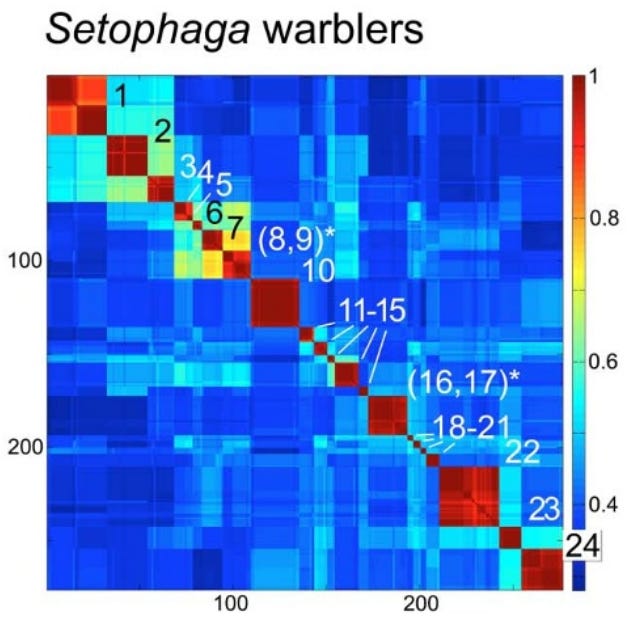
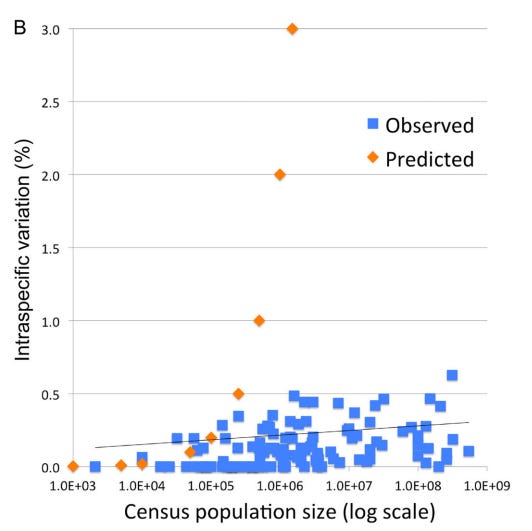
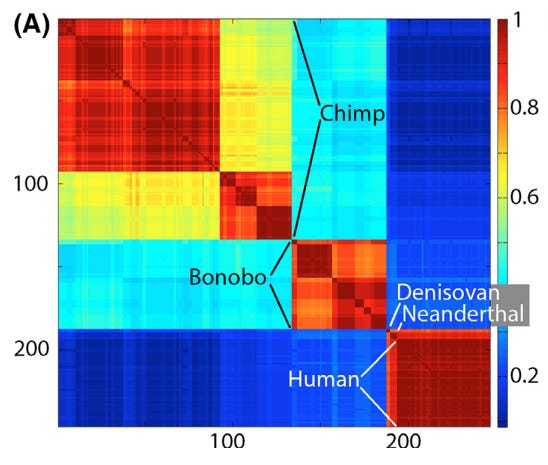
iNaturalist participation is unevenly distributed at the global, national, and local scale. I looked at county-level observations in Arizona to try to identify trends and potential causes of variability in total observations and percent of observations identified.
Introduction
Simple hypotheses for the total number of observations in
each county could be that the iNaturalist observations vary proportionally to
population (based on number of observers) or based on land area (based on availability
of organisms to observe). Alternately, a
hypothesis could be that areas with more species, or areas with more parks and
tourist attractions, might attract more attention from iNaturalist users.
Simple hypotheses for the percent of observations identified
could be that this varies with the population of a county, so that larger population
counties would be more likely to have more experts able to identify local
observations. Although counties with
more people might also have more total observations, so this hypothesis could
suggest that the percent of observations identified is either consistent across
counties or, if the identifier effect predominates, there could be a greater
percent in counties with more people.
Conversely, rural counties with low populations that also have lots of
observations from visitors (such as to national parks) might have a lower proportion
of observations identified.
I queried the iNaturalist.org website in December, 2025 for
total observations and Research Grade (RG) for each Arizona county. Some taxa, like fish or reptiles, might me
more abundant in one part of the state over another, so to ensure consistency,
I chose a taxa of species that is relatively evenly distributed across
Arizona: flowering plants
(angiosperms). I also obtained county
population and land area from publicly available information on the web. I then calculated % observations ID’d to RG,
total species observed, and total RG species confirmed for each county and
averages across all counties.
Results
Arizona has 15 counties that vary by orders of magnitude in
size, population, as well as other factors that contribute to iNaturalist
participation such as number of national parks and tourists. The largest county by population is Maricopa
county, with more than 4.5 million people, whereas the smallest county has less
than 10,000 people. The largest county
by land area is Coconino county, with more than 18,000 square miles, whereas
Santa Cruz, the smallest county, only has a little more than 1,000 square miles.
 |
| Figure 1 - Arizona Counties |
iNaturalist observations of flowering plants vary from only
about 3,000 in Greenlee county, to almost 300,000 in Pima county. RG observations vary from less than 2,000 in
Greenlee county, to more than 200,000 in Pima county, with corresponding
proportions of observations identified ranging from a low of 46% in Navajo
county to a high of 74% in La Paz county.
Counties vary in documented biodiversity from a high of
almost 1,900 species in Pima county, to a low of about 430 species in La Paz
county.
|
AZ
Counties |
angiosperm
observations |
RG
angiosperm |
% ID'd |
species
count |
RG
species |
% RG
species |
|
Pima |
298,000 |
211,000 |
71% |
2,396 |
1,855 |
77% |
|
Coconino |
134,440 |
61,500 |
46% |
2,197 |
1,489 |
68% |
|
Cochise |
122,500 |
79,550 |
65% |
1,860 |
1,460 |
78% |
|
Yavapai |
87,000 |
48,620 |
56% |
1,798 |
1,326 |
74% |
|
Santa Cruz |
47,000 |
28,000 |
60% |
1,586 |
1,273 |
80% |
|
Maricopa |
217,000 |
156,000 |
72% |
1,800 |
1,200 |
67% |
|
Pinal |
41,000 |
29,000 |
71% |
1,288 |
976 |
76% |
|
Gila |
26,233 |
15,500 |
59% |
1,240 |
938 |
76% |
|
Mohave |
36,000 |
26,000 |
72% |
1,200 |
890 |
74% |
|
Apache |
20,468 |
9,575 |
47% |
1,182 |
832 |
70% |
|
Graham |
12,700 |
7,780 |
61% |
1,044 |
774 |
74% |
|
Navajo |
14,000 |
6,400 |
46% |
1,000 |
714 |
71% |
|
Yuma |
18,800 |
13,800 |
73% |
682 |
521 |
76% |
|
Greenlee |
3,300 |
1,895 |
57% |
617 |
432 |
70% |
|
La Paz |
8,110 |
6,000 |
74% |
517 |
429 |
83% |
|
Average |
72,437 |
46,708 |
62% |
1,360 |
1,007 |
74% |
|
Total |
1,086,551 |
700,620 |
|
20,407 |
15,109 |
|
Table 1 Arizona Counties Angiosperm
plant observations and % Identified
To investigate trends across counties I used Tableau
Public to create graphs of total observations versus county land area and
population. Because Maricopa county has
3.5 million more people than the next largest county, I excluded it from the
analysis as an outlier.
While there is a trend toward more observations in larger
geographic areas, it is not very strong; the county with the greatest number of
observations is right in the middle of the distribution of sizes. The trend toward more observations with more
people is a bit better, but it is dominated by the 2nd most
populated county in Arizona, Pima county.
Removing that county from the analysis essentially eliminates the trend,
as the next largest county by population (Pinal) is below the average.

Figure 2 Observations by Land Area and Population, excluding Maricopa county.
I then looked at whether the activity of identifies varies
with land area or population, and again I did not find simple trends that
explain the variation. It does appear
that counties with smaller areas are more likely to have a greater proportion
of reported species and observations identified. This could be explained by the greater
tractability of small areas, where large counties have more possible habitats,
more potential species, and thus pose greater difficulty to botanists to
identify plants across the whole county.
While the largest county, Coconino, does have the largest number of
species reported, the smallest county (Santa Cruz) has more than the average
number of species reported. Also, two
counties that are in the middle of the distribution (Yavapai and Cochise), have
some of the highest number of reported species.

Figure 3 RG Observations %R by Land Area and Population, excluding Maricopa county.
The trend between population and proportion of identified
species is even weaker, with almost no trend apparent. This leads me to reject the hypothesis that
more people in an area necessarily lead to more identifications.
Instead, my current hypothesis, is that observation count is
driven by other factors outside of total population or land area. For example, I think that tourists who visit
natural wonders such as the Grand Canyon disproportionately increase the number
of observations and species in Coconino county, which results in a
corresponding decrease in the proportion of observations and species
identified. In contrast, a county that has
a strong resident population of people interested in nature, such as Pima county
(Tucson), has a large number of
observations and species, but also has a
large proportion of those observations identified to species.